Código
library(popdemo)
library(popbio)
library(ggplot2)
library(dplyr)
library(scales)
library(janitor)
library(tidyr)
library(flextable)Por: Demetria Mondragón, Ernesto Mujica Benitez y Elaine González
La elasticidad poblacional permite identificar qué transiciones del ciclo de vida tienen el mayor impacto proporcional sobre la tasa de crecimiento poblacional (\(\lambda\)). Este capítulo introduce la lógica, el cálculo y la interpretación aplicada de esta métrica.
La elasticidad describe cuánto cambia la tasa de crecimiento poblacional (\(\lambda\)) ante pequeñas variaciones proporcionales en los parámetros vitales de la matriz (supervivencia, crecimiento, fecundidad). A diferencia de la sensibilidad, que evalúa cambios absolutos, la elasticidad normaliza por la magnitud del parámetro y permite comparar la influencia relativa de transiciones con escalas distintas. En este módulo se muestra: (i) cómo derivar elasticidades a partir del modelo matricial de Leslie/Lefkovitch, (ii) por qué ciertas etapas (p. ej., supervivencia en adultos de larga vida) dominan \(\lambda\), y (iii) cómo interpretar estos resultados para priorizar acciones de manejo (protección de etapas clave, reducción de variabilidad en tasas vitales críticas, etc.). Incluimos ejemplos reproducibles con matrices reales y gráficos que contrastan elasticidades vs. sensibilidades, destacando cuándo la decisión de manejo cambiaría si usamos una u otra métrica. Finalizamos con recomendaciones prácticas: cómo reportar elasticidades con intervalos de confianza, cómo evitar interpretaciones erróneas cuando existen correlaciones entre transiciones, y cómo integrar elasticidades en un análisis LTRE o en simulaciones de escenarios. Esta lección se diferencia de las anteriores (fecundidad, crecimiento poblacional) porque conecta directamente la teoría con decisiones de conservación, mostrando el impacto marginal de cada transición sobre \(\lambda\) en términos comparables y accionables.
library(popdemo)
library(popbio)
library(ggplot2)
library(dplyr)
library(scales)
library(janitor)
library(tidyr)
library(flextable)source("R/rlt_theme.R") # tema + paleta canónicos (R/rlt_theme.R)Las matrices de elasticidad son un complemento de las matrices de transición que nos permiten evaluar el efecto de pequeños cambios proporcionales en los parámetros de la matriz sobre la tasa finita de crecimiento poblacional (\(\lambda\)); las entradas de la matriz de elasticidad con los valores más altos son las que más impacto tendrán sobre el comportamiento de \(\lambda\) (Kroon et al. 1986, Caswell 2001). Para matrices no negativas, las elasticidades de las entradas positivas son no negativas: aumentar \(a_{ij}\) incrementa (o no afecta) a \(\lambda\), y disminuirla la reduce (o no la afecta) (Caswell 2001). El uso de las elasticidades en ecología de plantas se remonta a 1974, cuando Sarukhán y Gadgil intentaron evaluar la importancia relativa de la reproducción sexual versus la reproducción vegetativa de tres especies de Ranunculus (Sarukhan y Gadgil 1974, Kroon et al. 1986).
El concepto de elasticidad en biología es un método orientado a evaluar el cambio proporcional en una variable en respuesta a un cambio proporcional en otra variable. En términos de matrices de transición, la elasticidad es una medida de la sensibilidad de la tasa de crecimiento poblacional a los cambios en alguna de las entradas de la matriz. En otras palabras, si uno cambia uno de los parámetros de la matriz de transición, la elasticidad mide cuánto cambiaría la tasa de crecimiento poblacional (Kroon et al. 1986). En biología, la elasticidad es una herramienta para entender la importancia del cambio de cada etapa de la historia de vida sobre la tasa de crecimiento poblacional, ya que permite identificar las etapas que más contribuyen a la tasa de crecimiento poblacional. Esta herramienta es muy útil para la conservación de especies y para la gestión de recursos naturales, porque en parte nos indica cuán sensible está el crecimiento poblacional cuando se cambian los parámetros de la historia de vida (crecimiento, supervivencia y fecundidad) de una especie. Un cambio proporcional se refiere a que, por ejemplo, si se cambió el crecimiento en 1%, qué efecto tuvo sobre el crecimiento intrínseco de la población, versus si se cambió el 1% de la fertilidad. De esta forma uno tiene una herramienta de comparación del impacto hipotético que tendrían los cambios, en el caso de que se pudieran lograr.
Los valores de elasticidad en la Biología de la Conservación se pueden interpretar como la vulnerabilidad de la población a perturbaciones en cada transición. Formalmente, la elasticidad cuantifica el efecto proporcional sobre \(\lambda\) de un cambio proporcional en una entrada de la matriz, suponiendo equilibrio demográfico y perturbaciones infinitesimales. Responde a la pregunta: ¿cuál de los cambios en las tasas vitales, tomados de uno en uno y manteniendo los demás constantes, produciría el mayor efecto sobre \(\lambda\)? En la práctica, las tasas vitales con una baja elasticidad suelen tener una alta variabilidad temporal, mientras que las de elevada elasticidad suelen variar poco — un patrón de canalización demográfica reportado por Ehrlén (1998). Por ello no solo se debe tener en cuenta cuán sensible es \(\lambda\) a una transición, sino también cuánta variación en esa transición ocurre realmente y cómo covaría con otras transiciones (Ehrlén y Van Groenendael 1998).
En términos matemáticos, la elasticidad es el cambio proporcional de \(\lambda\) ante un cambio proporcional en una entrada de la matriz. Concretamente, un cambio del 1% en \(a_{ij}\) produce aproximadamente \(e_{ij}\,\%\) de cambio en \(\lambda\) (válido para perturbaciones infinitesimales) (Kroon et al. 1986). La elasticidad \(e_{ij}\) de \(\lambda\) con respecto a \(a_{ij}\) (la entrada \(i, j\) de la matriz) se define como:
\[e_{ij}=\frac{a_{ij}}{\lambda}\,\frac{\partial \lambda}{\partial a_{ij}} = \frac{\partial \log \lambda}{\partial \log a_{ij}}\]
Es decir, \(e_{ij}\) es la pendiente de \(\log \lambda\) graficado contra \(\log a_{ij}\). Como los incrementos en escala logarítmica corresponden a cambios proporcionales en escala aritmética, la elasticidad mide la sensibilidad proporcional (en contraste con la sensibilidad absoluta \(\partial \lambda / \partial a_{ij}\)). Las elasticidades pueden organizarse en una matriz de elasticidad \(\mathbf{E}\) del mismo tamaño que la matriz de proyección:
\[\mathbf{E} = \frac{1}{\lambda}\,\mathbf{S} \circ \mathbf{A}\]
donde \(\mathbf{S}\) es la matriz de sensibilidad (con entradas \(s_{ij} = \partial \lambda / \partial a_{ij}\)), \(\mathbf{A}\) es la matriz de proyección y \(\circ\) es el producto de Hadamard (multiplicación elemento por elemento) (Caswell 2001). Aquí \(\lambda\) es la tasa finita de crecimiento (per-unidad-de-tiempo); su contraparte continua, la tasa intrínseca de incremento, es \(r = \log \lambda\).
Entradas de la matriz versus tasas vitales. Las elasticidades calculadas sobre \(a_{ij}\) son elasticidades de entradas de la matriz, no de las tasas vitales subyacentes. Una entrada suele ser el producto de varias tasas vitales: una transición \(a_{ij}\) típicamente combina supervivencia × probabilidad de progreso de estadio, y una entrada de fecundidad combina supervivencia × producción reproductiva per cápita. Por tanto, la elasticidad de \(a_{ij}\) y la elasticidad de cada tasa vital componente pueden diferir notablemente. Cuando una decisión de manejo se basa en una sola tasa vital (p. ej., supervivencia adulta), conviene descomponer la entrada y calcular la elasticidad a nivel de tasa vital (Franco y Silvertown 2004); ver también la sección de Limitaciones.
En contextos evolutivos, la elasticidad está estrechamente relacionada con el gradiente de selección sobre los componentes de la historia de vida: una entrada con alta elasticidad contribuye más a la aptitud poblacional, y por tanto está sujeta a selección más fuerte (Caswell 1989).
Las matrices de elasticidad tienen la ventaja de que la suma de las entradas es igual a uno, por lo que permite sumar diferentes entradas de la matriz, para evaluar la importancia de procesos (crecimiento, decrecimiento, permanencia o reproducción) o de estadios (plántulas, juveniles, adultos) sobre la \(\lambda\) (Silvertown et al. 1996). Esta cualidad ha favorecido que matrices de elasticidad sean una herramienta ampliamente utilizada para el desarrollo de planes de manejo y conservación de especies, ya que permite identificar a qué estadios o procesos se deben enfocar los esfuerzos de conservación (Tremblay 1997, Mondragón 2009, Berry y Cleavitt 2021, Timsina et al. 2021) o, en el caso opuesto, para determinar qué estadios pueden ser utilizados sin impactar fuertemente a la población (Mondragón 2009), es decir si quisiéramos aprovechar una especie, de qué estadio tendríamos que colectar, para impactar lo menos posible a la población.
La diferencia entre sensibilidad y elasticidad es que la sensibilidad mide el impacto en \(\lambda\) de cambiar la magnitud absoluta de una tasa vital particular en relación con el cambio de otras tasas vitales (Caswell 1989), lo que implica que los cambios en las tasas de crecimiento y permanencia tendrán un valor de uno (ya que estas tasas son proporciones), en tanto que los cambios en fecundidad podrán ser desde uno hasta cientos o miles, ya que la fecundidad se calcula con base en otra escala biológica (tal como número de semillas o plántulas). La elasticidad por su parte calcula la sensibilidad proporcional de \(\lambda\) a los cambios en una tasa vital (Kroon et al. 1986) evitando de esta forma el efecto de la magnitud del cambio. Cuanto mayor sea el valor de la elasticidad de la tasa vital, mayor será la contribución relativa de esa tasa vital al valor de lambda (\(\lambda\)).
Tomando como ejemplo la matriz de elasticidad de Erycina crista-galli una orquídea de ramilla (Mondragón et al. 2007), podremos recapitular todo lo previamente dicho (Tabla 11.1), así, lo primero que hay que recalcar es que el valor más alto de elasticidad para esta orquídea corresponde a la permanencia de los individuos adultos 2, es decir que cualquier pequeño cambio en el valor de dicha entrada (en la matriz de transición), tendrá el mayor impacto en el valor de lambda al comparar el valor de cambio en dicho parámetro cuando se modifiquen otras entradas de la matriz; segundo, es importante observar que al sumar todas las entradas de la matriz se obtiene un valor de uno. Lo mismo se mantiene cuando se suman todos los valores de elasticidad por estadio o por proceso. Es decir, si sumamos todos los valores de cada columna, tendremos el valor de elasticidad de cada estadio, para este caso el valor más alto lo ostenta el de adultos 2 (0.818), lo que implica que el destino de la población de E. crista-galli depende mayormente de lo que les pase a los individuos adultos.
etapas <- c("plántula", "juvenil", "adulto1", "adulto2")
ec <- matrix(
c(
0, 0, 0.004, 0.062,
0.195, 0, 0, 0,
0.171, 0.120, 0.053, 0.04,
0.073, 0.10, 0.435, 0.447
),
nrow = 4, ncol = 4, byrow = TRUE,
dimnames = list(etapas, etapas)
)
# ec# elas(ec)
# Convertir la matriz de elasticidad en un data frame
ec_df <- as.data.frame(elas(ec)) |> round(3)
# Sumar las elasticidades por etapas y añadir una fila con las sumas
ec_df <- ec_df %>%
bind_rows(summarise_all(., ~ if (is.numeric(.)) sum(.) else "Total"))ec_df <- ec_df %>%
mutate(stages = c("plántula", "juvenil", "adulto1", "adulto2", "suma")) %>% # Add the new column
relocate(stages, .before = everything())
ec_df %>%
flextable()stages | plántula | juvenil | adulto1 | adulto2 |
|---|---|---|---|---|
plántula | 0 | 0 | 0.001 | 0.058 |
juvenil | 0.015 | 0 | 0 | 0 |
adulto1 | 0.03 | 0.008 | 0.011 | 0.059 |
adulto2 | 0.014 | 0.007 | 0.096 | 0.702 |
suma | 0.059 | 0.015 | 0.108 | 0.819 |
Los valores anteriores son producidos por popbio::elas(), que internamente combina los autovectores derecho e izquierdo de la matriz con el producto de Hadamard. Para ilustrar de dónde sale un valor de elasticidad, podemos reconstruirlos paso a paso a partir de los autovectores. La sensibilidad se obtiene de:
\[s_{ij} = \frac{v_i\, w_j}{\langle v, w \rangle}\]
donde \(w\) es el autovector derecho dominante (estructura estable de estadios), \(v\) el autovector izquierdo dominante (valor reproductivo) y \(\langle v, w \rangle = \sum_k v_k w_k\). La matriz de elasticidad se obtiene como \(\mathbf{E} = (1/\lambda)\,\mathbf{S} \circ \mathbf{A}\) (Caswell 2001).
# Paso 1: autovalor dominante y autovectores
ev <- eigen(ec)
lambda <- Re(ev$values[1]) # tasa finita de crecimiento (lambda)
w <- Re(ev$vectors[, 1]) # autovector derecho: estructura estable
ev_t <- eigen(t(ec))
v <- Re(ev_t$vectors[, 1]) # autovector izquierdo: valor reproductivo
# Paso 2: matriz de sensibilidad S
S <- outer(v, w) / sum(v * w)
# Paso 3: matriz de elasticidad E = (1/lambda) * S o A
E_manual <- (S * ec) / lambda
round(E_manual, 3) plántula juvenil adulto1 adulto2
plántula 0.000 0.000 0.001 0.058
juvenil 0.015 0.000 0.000 0.000
adulto1 0.030 0.008 0.011 0.059
adulto2 0.014 0.007 0.096 0.702# Verificación: las entradas deben sumar a 1
sum(E_manual)[1] 1El resultado coincide con la Tabla 11.1 (salvo redondeo) y la suma de todas las entradas es 1, una propiedad fundamental de las matrices de elasticidad para matrices no negativas y primitivas (Kroon et al. 1986).
En la Tabla 11.2 se ven los diferentes valores de elasticidad por procesos en Erycina crista-galli (Mondragón et al. 2007) y el impacto por diferentes procesos de la historia de vida de la población. Ahora, si lo que quisiéramos saber es que proceso o tasa vital es lo que rige el comportamiento de la población, lo que se hace es sumar las entradas de la matriz que lo representan, en este caso hemos puesto en diferentes colores las entradas que manifiestan cada tasa, en rojo es la permanencia, en azul el crecimiento, en amarillo el decrecimiento o retrogresión y en verde la fecundidad. Eso nos permite determinar cuál es el impacto de elasticidad por procesos y tasas vitales, sumando cada uno de los valores en cada categoría. Es claro que el impacto principal para la población de E. crista-galli es la permanencia de los individuos, seguido por el crecimiento, en tanto que la fecundidad y la retrogresión tienen un impacto menor.
La suma de cada grupo:
# Matriz de elasticidad como data frame, con una columna que rotula la etapa
# de cada fila (la etapa de origen está en el nombre de cada columna).
ec_df <- as.data.frame(elas(ec)) |> round(3)
ec_df <- data.frame(Etapa = rownames(ec_df), ec_df, check.names = FALSE)
# Colores didácticos por tasa vital, siguiendo el texto: rojo = permanencia,
# azul = crecimiento, amarillo = retrogresión (decrecimiento), verde =
# fecundidad. Se usan tonos pastel para que las cifras en negro sigan legibles.
c_perm <- "#FBB4AE" # rojo claro — permanencia (diagonal)
c_crec <- "#B3CDE3" # azul claro — crecimiento (progresión)
c_retr <- "#FFFFCC" # amarillo — retrogresión (decrecimiento)
c_fec <- "#CCEBC5" # verde claro — fecundidad (reproducción)
ft <- flextable(ec_df)
# Permanencia: individuos que se quedan en su etapa (diagonal con valor > 0).
ft <- bg(ft, i = 3, j = "adulto1", bg = c_perm)
ft <- bg(ft, i = 4, j = "adulto2", bg = c_perm)
# Crecimiento: transición a una etapa posterior (debajo de la diagonal).
ft <- bg(ft, i = 2, j = "plántula", bg = c_crec)
ft <- bg(ft, i = 3, j = "plántula", bg = c_crec)
ft <- bg(ft, i = 3, j = "juvenil", bg = c_crec)
ft <- bg(ft, i = 4, j = "plántula", bg = c_crec)
ft <- bg(ft, i = 4, j = "juvenil", bg = c_crec)
ft <- bg(ft, i = 4, j = "adulto1", bg = c_crec)
# Retrogresión: regreso a una etapa anterior (adulto2 -> adulto1).
ft <- bg(ft, i = 3, j = "adulto2", bg = c_retr)
# Fecundidad: reproducción de adultos hacia plántulas (fila plántula).
ft <- bg(ft, i = 1, j = "adulto1", bg = c_fec)
ft <- bg(ft, i = 1, j = "adulto2", bg = c_fec)
ftEtapa | plántula | juvenil | adulto1 | adulto2 |
|---|---|---|---|---|
plántula | 0 | 0 | 0.001 | 0.058 |
juvenil | 0.015 | 0 | 0 | 0 |
adulto1 | 0.03 | 0.008 | 0.011 | 0.059 |
adulto2 | 0.014 | 0.007 | 0.096 | 0.702 |
Por lo que, de manera simplista, pudiéramos decir que, si quisiéramos desarrollar un plan de conservación para esta especie, nuestros esfuerzos debieran enfocarse en asegurar la supervivencia de los individuos adultos, ya que la mortandad (por causas naturales o por cosecha) de los individuos adultos tiene el mayor impacto sobre la persistencia de la población. En tanto que si quisiéramos hacer un uso sustentable de esta orquídea lo recomendable sería recolectar individuos juveniles, cuyos valores de elasticidad son menores a la de los adultos. El impacto de remover plántulas o juveniles de la población tendría menos impacto a largo plazo sobre la supervivencia de la población.
Importante: estas conclusiones suponen perturbaciones pequeñas. La elasticidad es una aproximación local que no captura efectos no lineales, compensación demográfica ni dependencia de la densidad; por ello no debe usarse para justificar cosechas sustanciales sin análisis adicionales (ver §Limitaciones y el capítulo de funciones de transferencia).
Las elasticidades también han sido utilizadas para establecer el comportamiento demográfico de las especies, lo que ha permitido generar patrones entre formas de vida, así como comparaciones tanto entre formas de vida como entre especies (Silvertown et al. 1996, Mondragón Chaparro et al. 2015). Así, por ejemplo, si quisiéramos comparar el comportamiento demográfico de las orquídeas de ramillas versus las orquídeas epífitas convencionales (Tabla 11.3) (reconstruido de (Mondragón et al. 2007)), veríamos que para ambos tipos de orquídeas lo más importante es la permanencia (supervivencia) de los individuos, en tanto que se puede observar una diferencia entre la orquídea de ramilla y las otras, en la contribución de los estadios al comportamiento de lambda, ya que para la primera, los individuos reproductivos presentan mayores valores de elasticidad (al igual que para Lepanthes eltoroensis), en tanto que para las otras orquídea los valores más altos de elasticidad los tienen los individuos no reproductivos (Mondragón et al. 2007).
En la Figura 11.4 se observan los valores de elasticidad añadidos por proceso de historia de vida para cinco especies de orquídeas epífitas:

Se observa que los valores de elasticidad más altos corresponden a la permanencia seguidos por el crecimiento.
Especie | Fecundidad | Estasis | Crecimiento | Plántulas | No_reproductivos | Reproductivos |
|---|---|---|---|---|---|---|
Erycina crista-galli | 0.035 | 0.718 | 0.247 | 0.0345 | 0.0456 | 0.9197 |
Artorima erubescens | 0 | 0.646 | 0.354 | 0.0023 | 0.7207 | 0.277 |
Laelia speciosa | 0.116 | 0.651 | 0.328 | 0.12 | 0.669 | 0.216 |
Lepanthes eltoroensis | 0.005 | 0.88 | 0.115 | 0.1133 | 0.0784 | 0.8083 |
Lepanthes caritensis | 0.002 | 0.885 | 0.114 | 0.018 | 0.648 | 0.335 |


EE <- Elasticidad_s %>%
select(Especie, Fecundidad, Estasis, Crecimiento) %>%
pivot_longer(
cols = -Especie,
names_to = "s",
values_to = "Elasticidad"
)
EE$s <- factor(
EE$s,
levels = c("Fecundidad", "Estasis", "Crecimiento")
)
ggplot(EE, aes(x = s, y = Elasticidad, fill = Especie)) +
geom_bar(stat = "identity", position = "dodge", color = "white") +
theme(
axis.text.x = element_text(angle = 45, hjust = 1),
legend.text = element_text(face = "italic")
) +
labs(
title = "Elasticidad por proceso demográfico",
x = "s",
y = "Elasticidad"
) +
rlt_style_fill()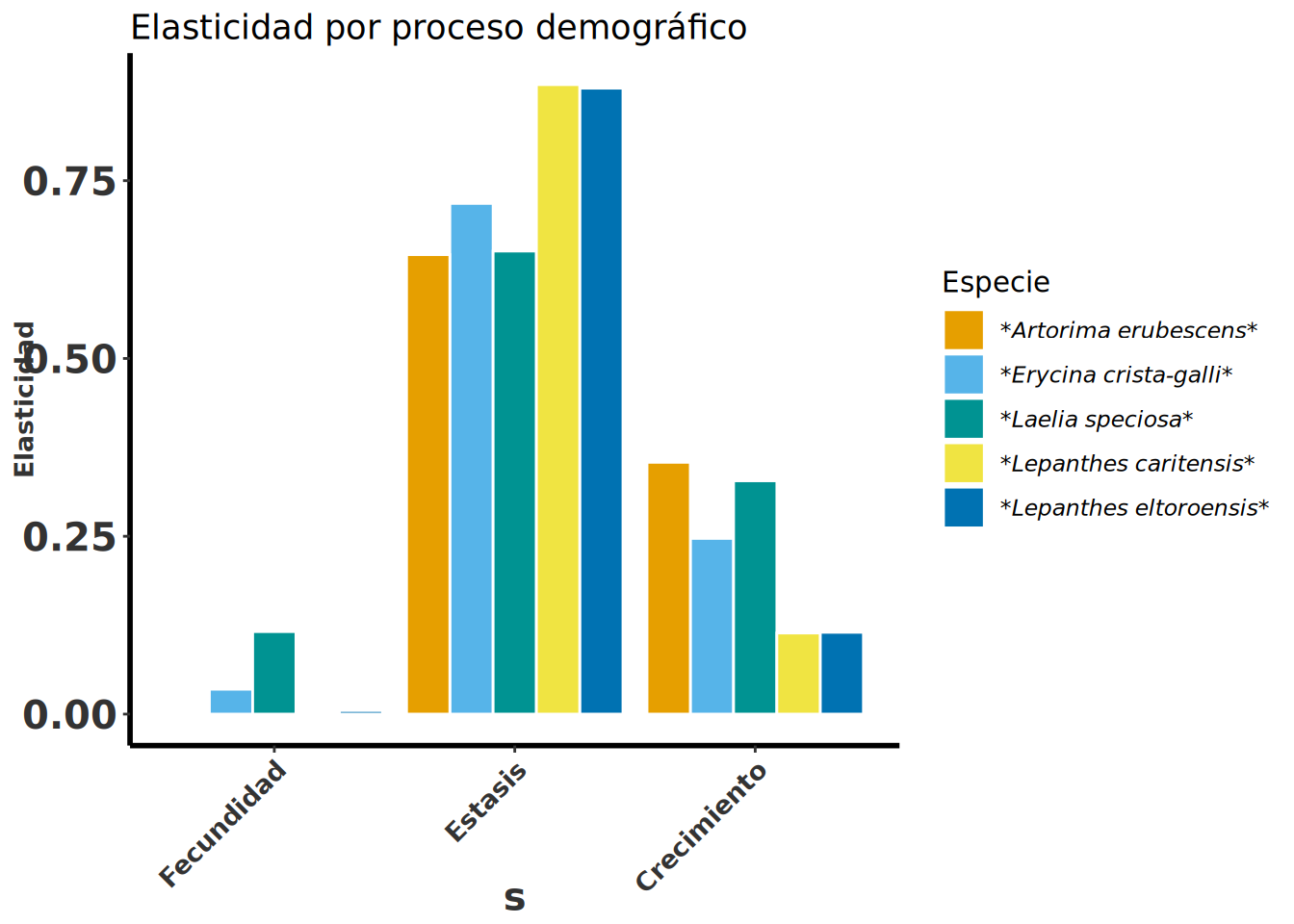
En la Figura 11.5 se observan los valores de elasticidad agrupados por estadios (plántulas, individuos no reproductivos y reproductivos). Los valores de elasticidad son más altos en los estadios no reproductivo en tres especies y reproductivos (adultos activos) en dos especies. Las plántulas tienen las elasticidad más pequeñas en todas las especies.
EE1 <- Elasticidad_s %>%
select(Especie, Plántulas, No_reproductivos, Reproductivos) %>%
pivot_longer(
cols = -Especie,
names_to = "s",
values_to = "Elasticidad"
)
EE1$s2 <- factor(
EE1$s,
levels = c("Plántulas", "No_reproductivos", "Reproductivos")
)
ggplot(EE1, aes(x = s2, y = Elasticidad, fill = Especie)) +
geom_bar(stat = "identity", position = "dodge", color = "white") +
theme(
axis.text.x = element_text(angle = 45, hjust = 1),
legend.text = element_text(face = "italic")
) +
labs(
title = "Elasticidad por proceso demográfico",
x = "Etapas",
y = "Elasticidad"
) +
rlt_style_fill()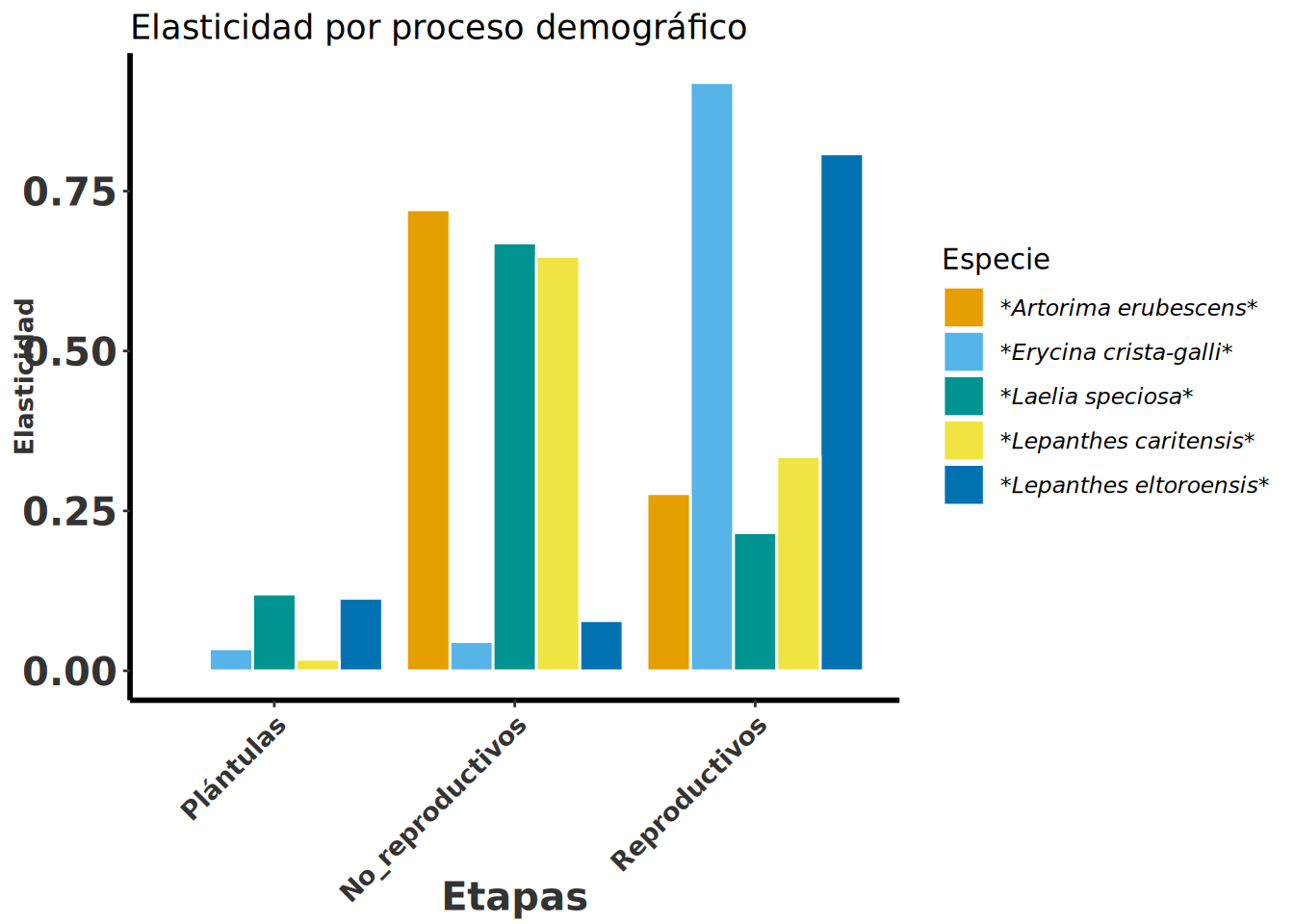
La elasticidad es una herramienta prospectiva: pregunta qué pasaría con \(\lambda\) si modificáramos hipotéticamente una entrada de la matriz, manteniendo todo lo demás constante. Su contraparte natural es el LTRE (Life Table Response Experiment), una herramienta retrospectiva que descompone diferencias observadas en \(\lambda\) entre poblaciones, tratamientos o periodos en contribuciones específicas de cada parámetro vital.
Las dos herramientas son complementarias: la elasticidad identifica las transiciones donde un cambio tendría mayor impacto potencial, y el LTRE cuantifica qué cambios efectivamente ocurrieron y cuánto contribuyeron al cambio observado en \(\lambda\). Por ejemplo, una transición puede tener alta elasticidad y, sin embargo, contribuir poco a una diferencia observada entre años porque varía poco entre años; el LTRE permite distinguir esto. Para la presentación formal del LTRE y un ejemplo aplicado, ver el capítulo Experimento de Respuesta de Tabla de Vida (Caswell 2001).

Aunque las matrices de elasticidad son una herramienta muy útil para determinar el comportamiento demográfico de una especie o un conjunto de estas, es necesario estar conscientes de que las diferentes entradas de dichas matrices son el resultado, en la mayoría de los casos, de la conjunción de varias tasas vitales a la vez (Franco y Silvertown 2004), además de que dependen fuertemente de la calidad de los datos. Así mismo, es importante considerar que ciertas entradas son la combinación de varios procesos demográficos. Por ejemplo, la entrada que representa la transición de plántula a juvenil implica que el individuo debe sobrevivir y además crecer lo suficiente para pasar al siguiente estadio; o en el caso de la fecundidad, que el individuo debe sobrevivir y además reproducirse. Por ello se recomienda separar los valores de cada proceso de cada entrada y calcular las elasticidades de las tasas vitales per se. Por consecuencia, el acercamiento más reciente de transient function desarrollado por Stott et al. (2012) pudiese ser un acercamiento más adecuado si los cambios son grandes, ya que muchos de los efectos no son lineales. Adicionalmente, si la población está lejos de su estructura estable, la elasticidad no es una buena aproximación de la respuesta de la población a los cambios en las tasas vitales (Stott et al. 2012). Por lo que es recomendable usar el análisis de elasticidad solamente para poblaciones que están cerca de su equilibrio y cuando los cambios en las tasas vitales son pequeños.
Una de las limitaciones principales del análisis de elasticidad tradicional mencionado arriba es que el modelo de respuestas es lineal. En otras palabras, se asume que los cambios en las tasas vitales son proporcionales a los cambios en la tasa de crecimiento poblacional y que, independientemente del tamaño del cambio, la respuesta seguirá un patrón lineal. En la realidad, los cambios en las tasas vitales no son proporcionales ni lineales a los cambios en la tasa de crecimiento poblacional. Por ejemplo, si uno aumenta mucho la supervivencia de los individuos de una etapa, la tasa de crecimiento poblacional no aumentará de manera proporcional y lineal. Por consecuencia, la elasticidad es una aproximación y no una descripción exacta de la relación entre los cambios en las tasas vitales y la tasa de crecimiento poblacional, y es aplicable solamente a análisis de perturbaciones pequeñas. Vea el capítulo de funciones de transferencia para métodos más avanzados de análisis de elasticidad.
Por último, las elasticidades calculadas mediante eigen.analysis() o popbio::elas() son deterministas: suponen un ambiente constante y una matriz fija. Cuando las tasas vitales varían entre años por variabilidad ambiental, conviene calcular elasticidades estocásticas sobre la tasa estocástica de crecimiento \(\log \lambda_s\) (Caswell 2001). Las elasticidades estocásticas pueden diferir notablemente de las deterministas, especialmente para tasas vitales con alta varianza temporal, y son la métrica adecuada cuando el ambiente es marcadamente variable. De forma análoga, dentro de un marco bayesiano la incertidumbre en los parámetros vitales se propaga directamente a las elasticidades, produciendo distribuciones posteriores en lugar de valores puntuales (ver el capítulo Acercamiento bayesiano para calcular transiciones y fecundidades).
etapas <- c("plántula", "juvenil", "adulto1", "adulto2")Capturar los datos de la matriz para Erycina crista-galli (Mondragón et al. 2007) (tabla 4 dentro de dicho artículo)
ec <- matrix(
c(
0, 0, 0.004, 0.062,
0.195, 0, 0, 0,
0.171, 0.120, 0.053, 0.04,
0.073, 0.10, 0.435, 0.447
),
nrow = 4, ncol = 4, byrow = TRUE,
dimnames = list(etapas, etapas)
)
ec plántula juvenil adulto1 adulto2
plántula 0.000 0.00 0.004 0.062
juvenil 0.195 0.00 0.000 0.000
adulto1 0.171 0.12 0.053 0.040
adulto2 0.073 0.10 0.435 0.447Introducir los valores iniciales por estadio (vector N): Cuantos individuos hay en cada estadio al inicio del análisis.
N0_ec <- matrix(c(41, 50, 246, 226), ncol = 1)
N0_ec [,1]
[1,] 41
[2,] 50
[3,] 246
[4,] 226eigen.analysis()Para calcular los autovalores y autovectores de la matriz se usa la función eigen.analysis() del paquete popbio. Se obtiene el crecimiento intrínseco Lambda, la sensibilidad, elasticidad, valor reproductivo y damping ratio. En el capítulo de propiedades se presenta la interpretación de los valores reproductivos y el damping ratio.
eigen.analysis(ec, zero = FALSE) # la función es del paquete popbio$lambda1
[1] 0.5214416
$stable.stage
plántula juvenil adulto1 adulto2
0.09190164 0.03436784 0.10775779 0.76597273
$sensitivities
plántula juvenil adulto1 adulto2
plántula 0.05877008 0.02197785 0.0689099 0.4898310
juvenil 0.03994690 0.01493867 0.0468391 0.3329455
adulto1 0.09172220 0.03430073 0.1075474 0.7644772
adulto2 0.09823314 0.03673558 0.1151817 0.8187439
$elasticities
plántula juvenil adulto1 adulto2
plántula 0.00000000 0.000000000 0.0005286107 0.05824146
juvenil 0.01493867 0.000000000 0.0000000000 0.00000000
adulto1 0.03007910 0.007893670 0.0109312556 0.05864336
adulto2 0.01375230 0.007045004 0.0960875206 0.70185904
$repro.value
plántula juvenil adulto1 adulto2
1.0000000 0.6797151 1.5606957 1.6714823
$damping.ratio
[1] 4.222528Tres direcciones complementarias profundizan los temas de este capítulo: